
Лекция 3. Репликация различных ДНК и ее регуляция и репарация
Уотсон и Крик предположили, что для удвоения ДНК должны произойти разрыв водородных связей, удерживающих вместе спиральный дуплекс, и расхождение цепей. Они также высказали мысль, что каждая цепь дуплекса служит матрицей при синтезе комплиментарной цепи и в результате образуются две пары цепей, в каждой из которых только одна является родительской. Уотсон и Крик предполагали, что репликация ДНК осуществляется спонтанно, без участия ферментов, но это оказалось неверно. Тем не менее идея о том, что удвоение ДНК происходит путем последовательного соединения нуклеотидов в соответствии с правилом комплиментарности, заданным каждой цепью спирали, разрешила концептуальную проблему точного воспроизведения генов.
С того времени как было высказано это предположение, матричная природа механизма репликации была подтверждена многочисленными данными, полученными как in vitro так и in vivo для различных организмов. Согласно модели репликация всех двуцепочечных ДНК полуконсервативна. Доказательство полуконсервативного механизма было получено в 1958 г. учеными Мезельсоном и Сталь(ем).
Сначала они выращивали бактерий длительное время на среде, содержащей тяжелый изотоп азота ( 15 N), который включался в ДНК, а затем переносили их на среду, содержащую обычный легкий изотоп азота (14 N).
После репликации дочернюю ДНК первого поколения фракционировали по плотности. Оказалось, что вся дочерняя ДНК однородна и имеет плотность, промежуточную между плотностью тяжелой и легкой ДНК. Следовательно, одна цепь молекулы дочерней ДНК содержала 15 N, а другая 14 N, что отвечает полуконсервативному механизму. Существуют ли в природе альтернативные способы репликации двуцепочечной ДНК (консервативный и дисперсный) – неизвестно. Итак, после одно раунда репликации одна цепь в каждой из двух дочерних ДНК является родительской, т.е. консервативной, а другая – синтезированной заново.
Репликация одноцепочечной ДНК у вирусов. Если геном представлен одноцепочечной ДНК (как у некоторых вирусов), то эта единственная цепь служит матрицей для образования комплиментарной цепи, с которой она образует дуплекс, а затем на этом дуплексе синтезируются либо дочерние дуплексы, либо одноцепочечные копии одной их матричных цепей. Репликация генетического материала вируса осуществляет обычно с участием ферментов клетки-хозяина. На некоторых молекулах вирусной ДНК синтезируются также ее ДНК-копии – с помощью либо клеточной, либо кодируемой вирусом ДНК-полимеразы. Эти ДНК-копии используются в последствии при сборке вирусных частиц. Репликация ДНК вирусов происходит либо в ядре клетки-хозяина (вирус герпеса), либо в цитоплазме (поксвирусы).
Методы определения нуклеиновой последовательности ДНК
... достоверности секвенирования; Выделить новейшие методы определения последовательности ДНК. 1. Нуклеиновые кислоты. Структура дезоксирибонуклеиновой кислоты (ДНК) Прежде чем приступить к анализу методов секвенирования, необходимо определить ... 10 пар нуклеотидов. Молекулы ДНК расположены 5'-конец одной цепи и 3'-конец другой цепи [25]. Все основания цепей ДНК расположены внутри двойной спирали, ...
Редупликацию ДНК
Теперь перейдем к добавлениям и уточнениям. Начнем с того, что в клетке кишечной палочки (E.coli) обнаружилась не одна, а целых три ДНК-полимеразы. Они заметно отличаются друг от друга по молекулярному весу и по числу молекул каждой из них, содержащихся в клетке. А также по их роли в процессе редупликации ДНК.
ДНК-полимераза I
«топоизомеразы».
Структура точек начала репликации., ДНК-геликаза,
Только теперь, сразу за праймером на ту же нить ДНК (условимся, для простоты, называть ее «первой») садится ДНК-полимераза, которая может начать строительство комплементарной нити ДНК только начиная от праймера, присоединяясь к нему («танцует от печки»).
Это — ДНК-полимераза III, самая крупная, состоящая из 6-ти субъединиц и по своей функции главная — она будет вести «комплементарный синтез» ДНК по этой первой материнской нити ДНК до самого конца. Первоначальное движение этой ДНК-полимеразы ограничено 1-2-мя тысячами нуклеотидов первой нити (у эукариотов — всего на 200 нуклеотидов).
Вторая материнская нить (еще пустая) формирует вместе с первой нитью «вилку редупликации».
«ДНК-связывающих белка».
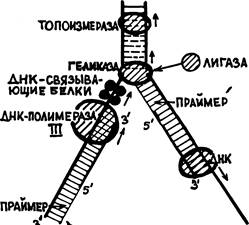
Пройдя до вершины вилки разошедшихся нитей материнской ДНК тандем геликаза-ДНК-связывающие белки — ДНК-полимераза III останавливается (см. рис. 28).
Топоизомераза уходит дальше по двухнитевой материнской ДНК, а геликаза разрывает сахаро-фосфатную связь на 2-й нити. Уплотненные на участке, прилегающем к вилке, витки двойной спирали расправляются, 1-я нить ДНК вместе с сидящими на ней белками вращается вокруг своей оси, а вокруг этой нити поворачивается и отрезанный кусок 2-й нити, временно связанный с геликазой. Этот кусок называют «фрагментом Оказаки» — по имени ученого, обнаружившего появление таких фрагментов при редупликации. После снятия напряжения нити двойной спирали материнской ДНК снова могут начать расходиться. Но до этого с отрезанного конца фрагмента Оказаки другая праймаза начинает на нем построение нового рибонуклеотидного праймера. Затем геликаза освобождает фрагмент и уходит вперед, а специальный фермент «лигаза» пришивает начало фрагмента Оказаки на прежнее место — ко 2-й нити материнской ДНК. Заметим, что лигаза (М=96 тыс.) в клетке E.coli представлена наиболее многочисленной популяцией — около 200 молекул. Из чего следует, что она выполняет не случайные «ремонтные» работы, а является полноправным членом совокупности ферментов, обеспечивающих редупликацию ДНК (подобно значению ниток для хирурга).
Когда праймер готов, впереди него, по направлению к 5′-концу 2-й материнской нити ДНК садится ДНК-полимераза I. Начинается строительство нити, комплементарной к этому фрагменту 2-й нити, опять в направлении 3’— 5′, считая по этой нити. ДНК-полимераза I доходит до конца фрагмента Оказаки и снимается. Этим заканчивается 1-й этап редупликации (рис. 28).
Роль наследственных факторов в формировании физического и психологического ...
... влиянии вредных факторов (в частности, алкоголя и курения) на развитие внутриутробного плода. Все эти сведения могут быть использованы для ранней диагностики и лечения наследственных заболеваний, профилактики врождённых пороков развития. ...
Между тем праймер, оставшийся в начале 1-й нити разрушается некой «рибонуклеазой Н», — ферментом, рвущим нить РНК, находящуюся в комплексе с нитью ДНК. На его место ДНК-полимераза II ставит «правильные» дезоксирибонуклеотиды. В то же время топоизомераза, геликаза, а за ними и ДНК-полимераза III продвигаются вперед.
Начинается 2-й этап редупликации. Вилка репликации тоже продвигается, прилегающий к ней участок материнской двунитевой ДНК уплотняется и весь синтезирующий тандем останавливается. Геликаза опять разрезает 2-ю нить, образуя второй фрагмент Оказаки. Так же, как раньше, на обрезанном (временно) конце фрагмента создается праймер, к нему «подсаживается» ДНК-полимераза I и начинает копировать второй фрагмент Оказаки, т.е. 2-ю нить материнской ДНК. Отличие второго этапа будет только в том, что на пути этой полимеразы встретится праймер, оставшийся от копирования 1-го фрагмента Оказаки. Но ДНК-полимераза I, в отличие от всех прочих ДНК-полимераз, обладает еще и 5’—3′ экзонуклеазной активностью, т. е. в направлении своего движения. Она разрушает праймер и доходит до того места, с которого начинала копирование 1-го фрагмента Оказаки ее предшественница. Остается только связать фосфодиэфирной связью эти два куска новосинтезированной комплементарной нити. Естественно, что это делает вездесущая ДНК-лигаза.
![]()
![]()
![]()
![]() рибонуклеаза Н
рибонуклеаза Н
ДНК-полимеразаII
Рис. 28
Тем временем в районе образования уже третьей вершины вилки редупликации происходят точно такие же события, как на 2-м этапе редупликации. Скорость этого процесса оценивается как, примерно, 1000 нуклеотидов в секунду у бактерий 100 — у животных и 20 — у растений.
Весьма вероятно, что в то же самое время аналогичные процессы расплетания двойной спирали с образованием фрагментов Оказаки и комплементарного построения новых нитей ДНК идут и с противоположного конца материнской ДНК. Разумеется, там ДНК-полимераза III непрерывно двигается вдоль той нити ДНК, которую мы назвали 2-й, а на фрагменты Оказаки разрезается 1-я нить. Когда два движения встречаются, две дочерние копии исходной ДНК оказываются готовы. (Их «сошьет» все та же лига-за.) Кстати оказалось, что длина фрагментов Оказаки у E.coli (1—2 тысячи нуклеотидов) значительно больше, чем у эукариотов (меньше 200).
Не лишено интереса совпадение этой последней цифры с длиной ДНК в нуклеосоме (см. ниже).
Более сложная модель движения репликативной вилки предполагает формирование реплисомы – мультиферментного комплекса более высокого уровня организации. Этот комплекс состоит из функционального праймосомо-праймазного комплекса, геликазы, полимеразы III, и, возможно, гиразы. Такой комплекс может обеспечивать удлинение лидирующей цепи и одновременно инициацию праймерной РНК, а также достраивание ДНК при синтезе отстающей цепи. Две реплисомы, работающие согласованно в двух вилках репликации, которые движутся в противоположных направлениях вдоль кольцевой хромосомы, сделали бы эту модель еще более изящной.
Пути, приёмы и средства улучшения памяти человека
... философской позиции. При написании реферата я использовала такие источники, как «Улучшаем память - в любом ... Классификация видов памяти память забывание запоминание По времени сохранения материала выделяются: o сенсорная память, осуществляющая сохранение ... Пути, приёмы и средства улучшения памяти Существует множество способов управления и тренировки памяти. Каждый человек хотел бы улучшить память, ...
Репликация кольцевых дуплексов. Репликация инициируется также в точке начала репликации (ori).
Растущие цепи образуют репликативные вилки, пересещающиеся либо в двух (вверху), либо в одном (внизу) направлении в зависимости от природы точки начала репликации.
В некоторых кольцевых геномах в каждой цепи имеется своя точка начала репликации (например, в митохондриальной ДНК животных ).
Синтез одной цепи начинается в точке ori R . Когда новая цепь доходит до точки oriD , начинается синтез другой цепи. Синтез инициируется путем образования праймерной РНК.
репликацией по типу катящегося кольца.
Что до механизма редупликации у эукариотов, то, хотя он изучен хуже, тем не менее и здесь найдены и охарактеризованы целых пять ДНК-полимераз, которые принято обозначать греческими буквами: α, β, γ, δ, и ε. Основным ферментом, подобным ДНК-полимеразе III у бактерий, является ДНК-полимераза δ. ДНК-полимераза α отвечает за построение праймеров (из рибонуклеотидов).
ДНК-полимераза β — копирует фрагменты Оказаки и отвечает за репарацию ДНК. ДНК-полимераза γ ведет синтез ДНК в митохондриях. Функция ДНК-полимеразы ε пока неизвестна.
Конечно, гигантские ДНК высших организмов начинают редупликацию не только с концов молекулы, но и во множестве промежуточных точек. Считают, что у дрожжей таких точек начала репликации около 300. Они отстоят друг от друга на 40 тысяч пар нуклеотидов. В ДНК человека насчитывают до 20 000 точек начала, расположенных с интервалом в 150 тысяч пар нуклеотидов. По-видимому, местами начала расплетания и посадки ДНК-полимеразы δ служат последовательности относительно слабо связанных А—Т пар оснований. После инициации репликация продолжается в двух направлениях от каждой точки до тех пор пока репликативные вилки двух соседних точек начала репликации не сольются. Полноразмерные ДНК каждой дочерней хромосомы получаются путем соединения более коротких независимо инициированных новосинтезированных цепей.
сателлитной ДНК
Строение центромер, Последовательности в области теломер.
Терминация репликации.
Терминация и расхождение в кольцевых геномах., Терминация и расхождение в линейных ДНК.
Первый : предполагает, что существуют цепи ДНК с прямыми повторами на концах (прозрачка 18).
После репликации два комплиментарных конца обоих незавершенных дуплексов могут спарится и образовать линейные конкатемеры с одноцепочечными разрывами.остающиеся пробелы могут быть заполнены путем удлинения цепей в направлении 3΄ →5΄ с последующим соединением их ДНК-лигазой либо путем прямого соединения стыкующихся концов с помощью ДНК-лигазы с образование конкатемеров. После надрезания конкатемера специфической эндонуклеазой образуются выступающие 5΄-концы, и ДНК-полимераза может наращивать более короткие цепи с 3΄-конца.
ГП курсовая. Основания приобретения права собственности Выполнила ...
... правовое регулирование процессов социального развития традиционно идет по пути регламентации отдельных оснований возникновения права собственности. Целью настоящей курсовой работы является определение оснований, способов приобретения права собственности, выявление их основных характеристики соотношения, также выявление пробелов ...
Второй. Предполагает наличие на конце каждой цепи ДНК коротких инвертированных повторов, благодаря которым образуются небольшие петли (прозрачка 19).
3΄-конец петли служит праймером для копирования нереплицированного участка. Благодаря специфическому разрыву в начале инвертированного повтора получается структура, которую можно достроить с 3΄-конца до восстановления исходной двуцепочечной концевой последовательности.
Отметим еще, что все ДНК-полимеразы, ведущие комплеметарный синтез, как у бактерий, так и у эукариотов, обладают еще и экзонуклеазной активностью в направлении 3′-5′, — обратном направлению синтеза. Они способны как бы «обернуться» и удалить только что ими же присоединенный нуклеотид. Это — очень важный механизм устранения ошибок в комплементарном синтезе. Ошибку ведь можно обнаружить только тогда, когда она уже сделана. И немедленно исправить! Это «умеют» делать ДНК-полимеразы. Такая коррекция не редкость, а норма. Считают, что без нее при редупликации ДНК из E.coli 5—10% нуклеотидов были бы присоединены неправильно. Благодаря коррекции 1 ошибка в этой ДНК приходится на десять миллионов пар оснований.
Репарация ДНК
ДНК – это единственная макромолекула клетки, которая способна устранять (репарировать) повреждения, возникающие в ее структуре. Более того, в ней закодирована информация о механизмах самых разнообразных репарационных процессов. Комплиментарное спаривание лежит в основе не только репликации ДНК, но процесса восстановления исходной структуры ДНК при репарации повреждений, затрагивающих остов молекулы, модификаций того или иного основания или ошибочного спаривания при рекомбинации (см. ниже).
Одновременное повреждение обеих цепей в одном месте и двуцепочечные разрывы часто оказываются летальными для ДНК, поскольку такие дефекты репарируются лишь в редких случаях.
Наиболее часто происходит разрыв гликозидных связей между пурином и дезоксирибозой N (депуринизация) при повышении температуры. За сутки в клетке человека совершается от 5000 до 10000 актов депуринизации -–это ведет к нарушению репликации и экспрессии генов. Кроме того, остатки Ц и А могут подвергаться спонтанному дезаминированию с образование соответственно остатков У и гипоксантина; частота таких событий примерно 100 на геном – это ведет к появлению мутаций.
Многие изменения в структуре ДНК происходят под действием химических веществ, присутствующих в окружающей среде. Это алкилирующие агенты (азотистые соединения, алкилсульфонаты, нитрозомочевина), которые модифицируют предпочтительно Г-остатки, соединения, встраивающиеся между соседними парами оснований и приводящие к появлению вставок и делеций во время репликации; бифункциональные агенты, способные образовывать ковалентные сшивки между двумя цепями ДНК и блокировать их расхождение при репликации. Не менее разрушительны физические воздействия – поглощение Т или Ц УФ-света приводит к образованию циклобутановых димеров между соседними пиримидинами; ионизирующая радиация (космические лучи) способствует образованию высокореакционоспособных свободных радикалов, оказывающих на ДНКсамые разнообразные воздействия; рентгеновские лучи – вызывают в ДНК одно-и-двуцепочечные разрывы, а также другие повреждения, характерные для воздействия свободных радикалов.
Известны два основных способа репарационных процессов:
Восстановление работе
... работников. Решение об удовлетворении иска о восстановлении на работе принимается в случае, если увольнение произведено без законного основания или с нарушением установленного порядка увольнения. ... признано незаконным, поскольку фактически отсутствовало основание, предусмотренное законом. Суд Шумилинского района рассмотрел дело по иску У. о восстановлении на работе. По делу установлено, что истец ...
- непосредственное исправление модификаций или неправильных спариваний, не требующее репликации для восстановления исходной структуры;
- удаление нуклеотидов, окружающих ошибочно спаренные или измененные пары оснований, и ресинтез этого участка путем репликации.
репарация путем прямого восстановления исходной структуры.
Если под воздействием алкилирующих агентов – N-метил –N-нитрозомочевины или N 1 , N-диметилнитрозогуанидина, в ДНК образовался О6 — метил- или О6 -алкилзамещенные гуаниновые остатки, то деалкилирование таких остатков идет при участии ферментов – О6 -метилгуанин-ДНК-алкилтрансферазы, которая катализирует перенос алкильных групп на сульфгидрильные группы цистеиновых остатков фермента, при этом акцепторный белок инактивируется. Это у бактерий и млекопитающих.
Если при облучении ДНК УФ-светом образовались циклобутановые димеры между соседними парами пиримидиновых оснований, то осуществляется ферментативное (фотолиазы –нет у млекопитающих) превращение их в мономеры при освещении раствора видимым светом в диапазоне длин волн 300-600 нм. Фермент образует стабильный комплекс с пиримидиновым димером и используя энергию поглощенног им света разрушает димер без разрыва цепей ДНК.
Репарация путем замены модифицированных остатков.
Замена модифицированного нуклеотида обычно проходит в четыре этапа:
1 этап . Фермент распознает этот нуклеотид и надрезает полинуклеотидную цепь вблизи него либо разрывает гликозидную связь между модифицированным основанием и дезоксирибозой. Выщепление сайтов, в которых произошла депуринизация или депиримидинизация осуществляется ферментами АР (апуриновые, апиримидиновые)-эндонуклеазы. В репарации N-алкилированных пуринов и других модифицированных оснований ключевую роль играют ферменты – N-гликозилазы, расщепляющие гликозидную связь между модифицированным основанием и дезоскирибозой (прозрачка 25)
2 этап . Экзонуклеаза удаляет модифицированный нуклеотид и/или соседние нуклеотиды, оставляя небольшую брешь.
3 этап . Удаленный участок синтезируется заново с 3΄-ОН-конца с использованием в качестве матрицы противоположной цепи.
4 этап . Концы разрыва, образовавшиеся в результате репарации, соединяются с восстановлением ковалентной целостности репарированной цепи (прозрачка 26)
Значение репарации ДНК.
Устранение ошибок репликации важно, так как большая часть повреждений блокирует передачу генетической информации последующему поколению, а остальные если их не устранить, сохраняться в геноме потомков и приведут к драматическим изменениям в молекулах белков, ферментов, необходимых для поддержания жизнедеятельности клетки. При повреждении определенных звеньев системы репарации клетки становятся особенно уязвимыми для некоторых химических и физических агентов. Люди, страдающие, например, пигментной ксеродермой, очень чувствительны к УФ-свету, и у них развиваются разные формы рака кожи даже при очень слабом воздействии солнечного света. Клетки таких людей несут мутацию в RAD-генах, проявляющуюся в том, что у них нарушена способность к выщеплению пиримидиновых димеров из УФ-облученной ДНК. Заболевание может быть обусловлено мутауцией в одном из по крайней мере девяти генов, что говорит о достаточно сложном механизме репарации ДНК, содержащей тиминовые димеры, у человека.как правило, заболевание бывает связано с неспособностью к выщеплению тиминовых димеров. Если к облученным клеткам в культуре добавить фермент, обладающий тиминдимергликозилазной и АР-эндонуклеазной активностями, то УФ-повреждения могут быть устранены.
Повреждение и гибель клеток и тканей
... физическими, химическими агентами. Ишемическое и гипоксическое повреждение; Повреждение, вызванное свободными радикалами, включая активированный кислород; Токсическое повреждение. Ишемическое и гипоксическое повреждение. Чаще обусловлено окклюзией артерий. Основными механизмами гибели клетки при гипоксии являются нарушение окислительного фосфорилирования, ...